The Krebs cycle is a series of enzymatic reactions that catalyzes the aerobic metabolism of fuel molecules to carbon dioxide and water, thereby generating energy for the production of adenosine triphosphate (ATP) molecules. The Krebs cycle is so named because much of its elucidation was the work of the British biochemist Hans Krebs. Many types of fuel molecules can be drawn into and utilized by the cycle, including acetyl coenzyme A (acetyl CoA), derived from glycolysis or fatty acid oxidation . Some amino acids are metabolized via the enzymatic reactions of the Krebs cycle. In eukaryotic cells , all but one of the enzymes catalyzing the reactions of the Krebs cycle are found in the mitochondrial matrixes.
The sequence of events known as the Krebs cycle is indeed a cycle; oxaloacetate is both the first reactant and the final product of the metabolic pathway (creating a loop). Because the Krebs cycle is responsible for the ultimate oxidation of metabolic intermediates produced during the metabolism of fats, proteins, and carbohydrates, it is the central mechanism for metabolism in the cell. In the first reaction of the cycle, acetyl CoA condenses with oxaloacetate to form citric acid. Acetyl CoA utilized in this way by the cycle has been produced either via the oxidation of fatty acids, the breakdown of certain amino acids, or the oxidative decarboxylation of pyruvate (a product of glycolysis). The citric acid produced by the condensation of acetyl CoA and oxaloacetate is a tricarboxylic acid containing three carboxylate groups. (Hence, the Krebs cycle is also referred to as the citric acid cycle or tricarboxylic acid cycle.)
After citrate has been formed, the cycle machinery continues through seven distinct enzyme-catalyzed reactions that produce, in order, isocitrate, α -ketoglutarate, succinyl coenzyme A, succinate, fumarate, malate, and
oxaloacetate. The freshly produced oxaloacetate, in turn, reacts with yet another molecule of acetyl CoA, and the cycle begins again. Each turn of the Krebs cycle produces two molecules of carbon dioxide, one guanosine triphosphate molecule (GTP), and enough electrons to generate three molecules of NADH and one molecule of FADH2.
The Krebs cycle is present in virtually all eukaryotic cells that contain mitochondria, but functions only as part of aerobic metabolism (when oxygen is available). This oxygen requirement is owing to the close relationship between the mitochondrial electron transport chain and the Krebs cycle. In the Krebs cycle, four oxidation–reduction reactions occur. A high energy phosphate bond in the form of GTP is also generated. (This high energy phosphate bond is later transferred to adenosine diphosphate [ADP] to form adenosine triphosphate [ATP].) As the enzymes of the Krebs cycle oxidize fuel molecules to carbon dioxide, the coenzymes NAD+, FAD, and coenzyme Q (also known as ubiquinone) are reduced. In order for the cycle to continue, these reduced coenzymes must become reoxidized by transferring their electrons to oxygen, thus producing water. Therefore, the final acceptor of the electrons produced by the oxidation of fuel molecules as part of the Krebs cycle is oxygen. In the absence of oxygen, the Krebs cycle is inhibited.
The citric acid cycle is an amphibolic pathway, meaning that it can be used for both the synthesis and degradation of biomolecules. Besides acetyl CoA (generated from glucose , fatty acids, or ketogenic amino acids), other biomolecules are metabolized by the cycle. Several amino acids are degraded to become what are intermediates of the cycle. Likewise, odd-chain fatty acids are metabolized to form succinyl coenzyme A, another intermediate of the cycle. Krebs cycle intermediates are also used by many organisms for the synthesis of other important biomolecules. Some organisms use the Krebs cycle intermediates α -ketoglutarate and oxaloacetate in the synthesis of several amino acids. Succinyl coenzyme A is utilized in the synthesis of porphyrin rings, used in heme manufacture and chlorophyll biosynthesis . Oxaloacetate and malate are utilized in the synthesis of glucose, in a process known as gluconeogenesis.
Lets break it down
The citric acid cycle
Overview and steps of the citric acid cycle, also known as the Krebs cycle or tricarboxylic acid (TCA) cycle.
Introduction
How important is the citric acid cycle? So important that it has not one, not two, but three different names in common usage today!
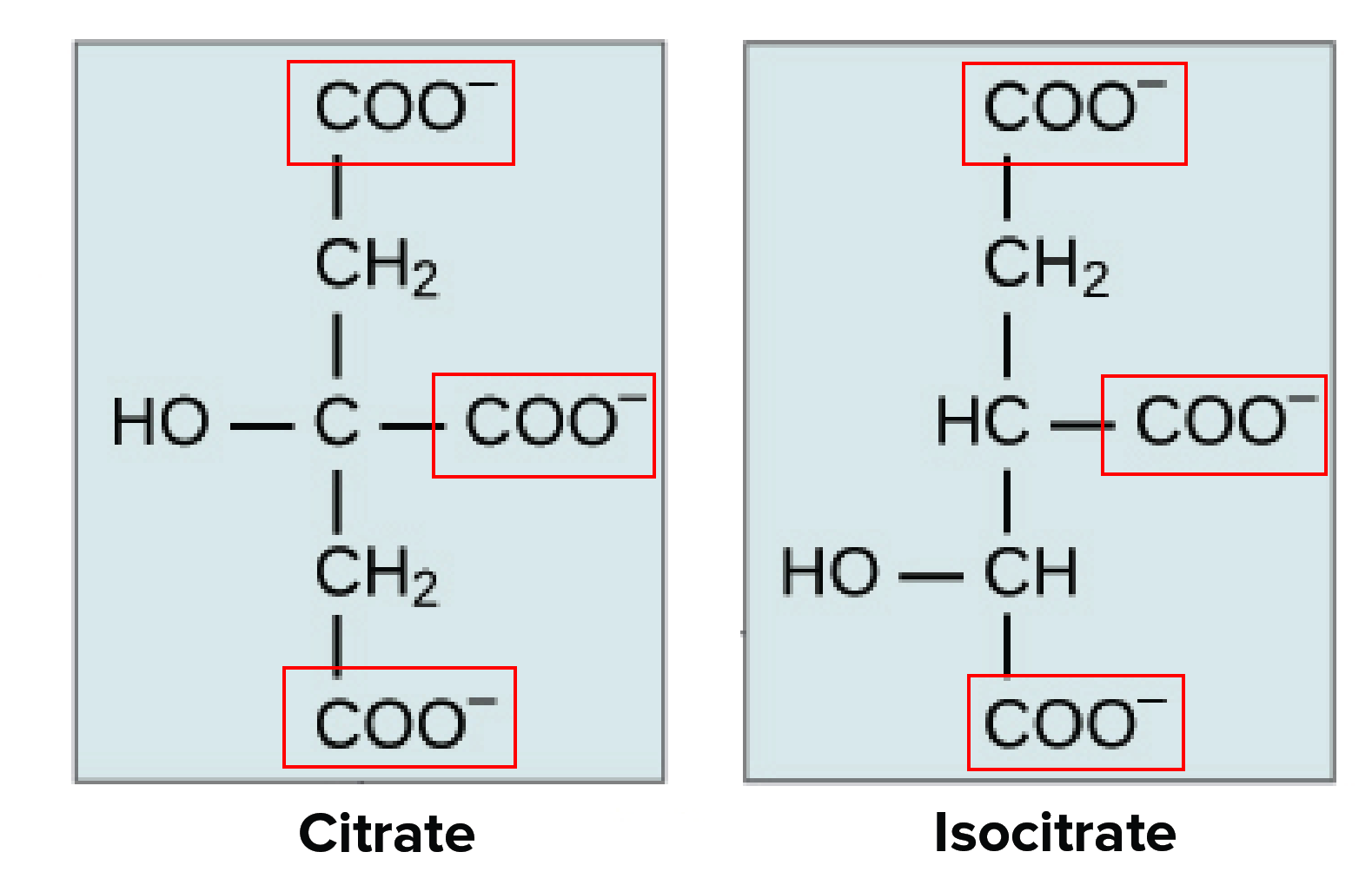
The name we’ll primarily use here, the citric acid cycle, refers to the first molecule that forms during the cycle’s reactions—citrate, or, in its protonated form, citric acid. However, you may also hear this series of reactions called the tricarboxylic acid (TCA) cycle, for the three carboxyl groups on its first two intermediates, or the Krebs cycle, after its discoverer, Hans Krebs.
The first two intermediates of the citric acid cycle are shown below. Each has three carboxyl groups, marked with red boxes. When citrate gains three H+\text {H}^+H+H, start superscript, plus, end superscript ions, so that it no longer has a negative charge, it is called citric acid.
Whatever you prefer to call it, the citric cycle is a central driver of cellular respiration. It takes acetyl CoA\text{CoA}CoAC, o, A—produced by the oxidation of pyruvate and originally derived from glucose—as its starting material and, in a series of redox reactions, harvests much of its bond energy in the form of NADH\text{NADH}NADHN, A, D, H, FADH2\text{FADH}_2FADH2F, A, D, H, start subscript, 2, end subscript, and ATP\text{ATP}ATPA, T, P molecules. The reduced electron carriers—NADH\text{NADH}NADHN, A, D, H and FADH2\text{FADH}_2FADH2F, A, D, H, start subscript, 2, end subscript—generated in the TCA cycle will pass their electrons into the electron transport chain and, through oxidative phosphorylation, will generate most of the ATP produced in cellular respiration.
Below, we’ll look in more detail at how this remarkable cycle works.
Overview of the citric acid cycle
In eukaryotes, the citric acid cycle takes place in the matrix of the mitochondria, just like the conversion of pyruvate to acetyl CoA\text{CoA}CoAC, o, A. In prokaryotes, these steps both take place in the cytoplasm. The citric acid cycle is a closed loop; the last part of the pathway reforms the molecule used in the first step. The cycle includes eight major steps.
Simplified diagram of the citric acid cycle. First, acetyl CoA combines with oxaloacetate, a four-carbon molecule, losing the CoA group and forming the six-carbon molecule citrate. After citrate undergoes a rearrangement step, it undergoes an oxidation reaction, transferring electrons to NAD+ to form NADH and releasing a molecule of carbon dioxide. The five-carbon molecule left behind then undergoes a second, similar reaction, transferring electrons to NAD+ to form NADH and releasing a carbon dioxide molecule.
The four-carbon molecule remaining then undergoes a series of transformations, in the course of which GDP and inorganic phosphate are converted into GTP—or, in some organisms, ADP and inorganic phosphate are converted into ATP—an FAD molecule is reduced to FADH2, and another NAD+ is reduced to NADH. At the end of this series of reactions, the four-carbon starting molecule, oxaloacetate, is regenerated, allowing the cycle to begin again.
In the first step of the cycle, acetyl CoA\text{CoA}CoAC, o, A combines with a four-carbon acceptor molecule, oxaloacetate, to form a six-carbon molecule called citrate. After a quick rearrangement, this six-carbon molecule releases two of its carbons as carbon dioxide molecules in a pair of similar reactions, producing a molecule of NADH\text{NADH}NADHN, A, D, H each time1^11start superscript, 1, end superscript. The enzymes that catalyze these reactions are key regulators of the citric acid cycle, speeding it up or slowing it down based on the cell’s energy needs2^22start superscript, 2, end superscript.
The remaining four-carbon molecule undergoes a series of additional reactions, first making an ATP\text{ATP}ATPA, T, P molecule—or, in some cells, a similar molecule called GTP\text{GTP}GTPG, T, P—then reducing the electron carrier FAD\text{FAD}FADF, A, D to FADH2\text{FADH}_2FADH2F, A, D, H, start subscript, 2, end subscript, and finally generating another NADH\text{NADH}NADHN, A, D, H. This set of reactions regenerates the starting molecule, oxaloacetate, so the cycle can repeat.
Overall, one turn of the citric acid cycle releases two carbon dioxide molecules and produces three NADH\text{NADH}NADHN, A, D, H, one FADH2\text{FADH}_2FADH2F, A, D, H, start subscript, 2, end subscript, and one ATP\text{ATP}ATPA, T, P or GTP\text{GTP}GTPG, T, P. The citric acid cycle goes around twice for each molecule of glucose that enters cellular respiration because there are two pyruvates—and thus, two acetyl CoA\text{CoA}CoAC, o, As—made per glucose.
Steps of the citric acid cycle
You’ve already gotten a preview of the molecules produced during the citric acid cycle. But how, exactly, are those molecules made? We’ll walk through the cycle step by step, seeing how NADH\text{NADH}NADHN, A, D, H, FADH2\text{FADH}_2FADH2F, A, D, H, start subscript, 2, end subscript, and ATP\text{ATP}ATPA, T, P/GTP\text{GTP}GTPG, T, P are produced and where carbon dioxide molecules are released.
Step 1. In the first step of the citric acid cycle, acetyl CoA\text{CoA}CoAC, o, A joins with a four-carbon molecule, oxaloacetate, releasing the CoA\text{CoA}CoAC, o, A group and forming a six-carbon molecule called citrate.
Step 2. In the second step, citrate is converted into its isomer, isocitrate. This is actually a two-step process, involving first the removal and then the addition of a water molecule, which is why the citric acid cycle is sometimes described as having nine steps—rather than the eight listed here3^33start superscript, 3, end superscript.
Step 3. In the third step, isocitrate is oxidized and releases a molecule of carbon dioxide, leaving behind a five-carbon molecule—α-ketoglutarate. During this step, NAD+\text{NAD}^+NAD+N, A, D, start superscript, plus, end superscript is reduced to form NADH\text{NADH}NADHN, A, D, H. The enzyme catalyzing this step, isocitrate dehydrogenase, is important in regulating the speed of the citric acid cycle.
Step 4. The fourth step is similar to the third. In this case, it’s α-ketoglutarate that’s oxidized, reducing NAD+\text{NAD}^+NAD+N, A, D, start superscript, plus, end superscript to NADH\text{NADH}NADHN, A, D, H and releasing a molecule of carbon dioxide in the process. The remaining four-carbon molecule picks up Coenzyme A, forming the unstable compound succinyl CoA\text{CoA}CoAC, o, A. The enzyme catalyzing this step, α-ketoglutarate dehydrogenase, is also important in regulation of the citric acid cycle.
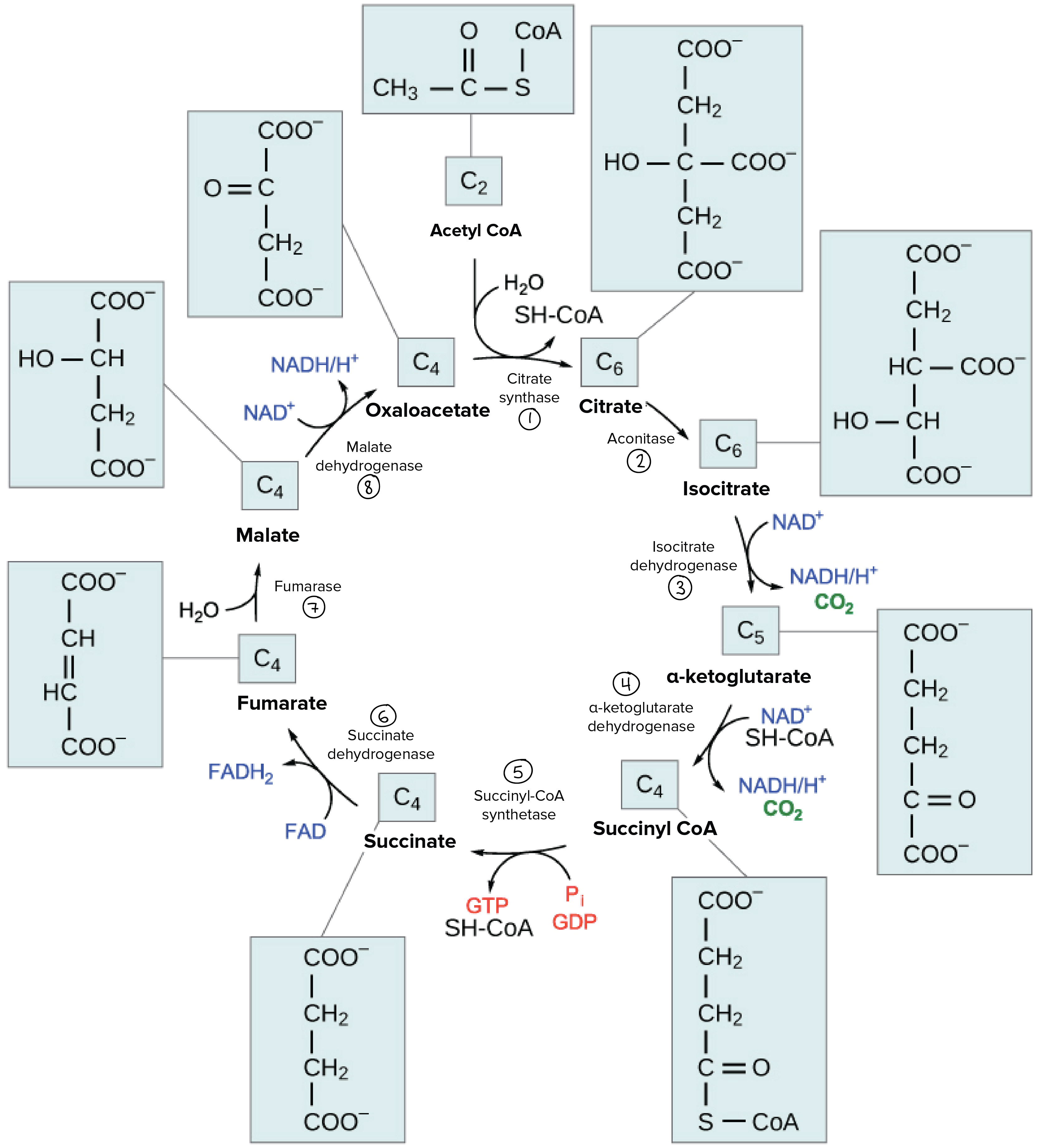
Detailed diagram of the citric acid cycle, showing the structures of the various cycle intermediates and the enzymes catalyzing each step.
Step 1. Acetyl CoA combines with oxaloacetate in a reaction catalyzed by citrate synthase. This reaction also takes a water molecule as a reactant, and it releases a SH-CoA molecule as a product.
Step 2. Citrate is converted into isocitrate in a reaction catalyzed by aconitase.
Step 3. Isocitrate is converted into α-ketoglutarate in a reaction catalyzed by isocitrate dehydrogenase. An NAD+ molecule is reduced to NADH + H+ in this reaction, and a carbon dioxide molecule is released as a product.
Step 4. α-ketoglutarate is converted to succinyl CoA in a reaction catalyzed by α-ketoglutarate dehydrogenase. An NAD+ molecule is reduced to NADH + H+ in this reaction, which also takes a SH-CoA molecule as reactant. A carbon dioxide molecule is released as a product.
Step 5. Succinyl CoA is converted to succinate in a reaction catalyzed by the enzyme succinyl-CoA synthetase. This reaction converts inorganic phosphate, Pi, and GDP to GTP and also releases a SH-CoA group.
Step 6. Succinate is converted to fumarate in a reaction catalyzed by succinate dehydrogenase. FAD is reduced to FADH2 in this reaction.
Step 7. Fumarate is converted to malate in a reaction catalyzed by the enzyme fumarase. This reaction requires a water molecule as a reactant.
Step 8. Malate is converted to oxaloacetate in a reaction catalyzed by malate dehydrogenase. This reaction reduces an NAD+ molecule to NADH + H+.
Step 5. In step five, the CoA\text{CoA}CoAC, o, A of succinyl CoA\text{CoA}CoAC, o, A is replaced by a phosphate group, which is then transferred to ADP\text{ADP}ADPA, D, P to make ATP\text{ATP}ATPA, T, P. In some cells, GDP\text{GDP}GDPG, D, P—guanine diphosphate—is used instead of ADP\text{ADP}ADPA, D, P, forming GTP\text{GTP}GTPG, T, P—guanine triphosphate—as a product. The four-carbon molecule produced in this step is called succinate.
GTP\text{GTP}GTPG, T, P is similar to ATP\text{ATP}ATPA, T, P: both serve as energy sources, and the two can be readily interconverted. Which of the two molecules is produced during the citric acid cycle depends on the organism and cell type. For example, ATP\text{ATP}ATPA, T, P is made in human heart cells, but GTP\text{GTP}GTPG, T, P is made in liver cells.
Step 6. In step six, succinate is oxidized, forming another four-carbon molecule called fumarate. In this reaction, two hydrogen atoms—with their electrons—are transferred to FAD\text{FAD}FADF, A, D, producing FADH2\text{FADH}_2FADH2F, A, D, H, start subscript, 2, end subscript. The enzyme that carries out this step is embedded in the inner membrane of the mitochondrion, so FADH2\text{FADH}_2FADH2F, A, D, H, start subscript, 2, end subscript can transfer its electrons directly into the electron transport chain.
Why use FAD here?
FAD\text{FAD}FADF, A, D is a better electron acceptor than NAD+\text{NAD}^+NAD+N, A, D, start superscript, plus, end superscript, meaning that it has a higher affinity, or “hunger”, for electrons. Succinate is not a great electron donor, meaning that it has a fairly high affinity for electrons itself and is not eager to give them up. NAD+\text{NAD}^+NAD+N, A, D, start superscript, plus, end superscript is not electron-hungry enough to pull electrons away from succinate, but FAD\text{FAD}FADF, A, D is4,5^{4,5}4,5start superscript, 4, comma, 5, end superscript.
Step 7. In step seven, water is added to the four-carbon molecule fumarate, converting it into another four-carbon molecule called malate.
Step 8. In the last step of the citric acid cycle, oxaloacetate—the starting four-carbon compound—is regenerated by oxidation of malate. Another molecule of NAD+\text{NAD}^+NAD+N, A, D, start superscript, plus, end superscript is reduced to NADH\text{NADH}NADHN, A, D, H in the process.
Products of the citric acid cycle
Let’s take a step back and do some accounting, tracing the fate of the carbons that enter the citric acid cycle and counting the reduced electron carriers—NADH\text{NADH}NADHN, A, D, H and FADH2\text{FADH}_2FADH2F, A, D, H, start subscript, 2, end subscript—and ATP\text{ATP}ATPA, T, P produced.
In a single turn of the cycle,
two carbons enter from acetyl CoA\text{CoA}CoAC, o, A, and two molecules of carbon dioxide are released;
three molecules of NADH\text{NADH}NADHN, A, D, H and one molecule of FADH2\text{FADH}_2FADH2F, A, D, H, start subscript, 2, end subscript are generated; and
one molecule of ATP\text{ATP}ATPA, T, P or GTP\text{GTP}GTPG, T, P is produced.
These figures are for one turn of the cycle, corresponding to one molecule of acetyl CoA\text{CoA}CoAC, o, A. Each glucose produces two acetyl CoA\text{CoA}CoAC, o, A molecules, so we need to multiply these numbers by 2222 if we want the per-glucose yield.
Two carbons—from acetyl CoA\text{CoA}CoAC, o, A—enter the citric acid cycle in each turn, and two carbon dioxide molecules are released. However, the carbon dioxide molecules don’t actually contain carbon atoms from the acetyl CoA\text{CoA}CoAC, o, A that just entered the cycle. Instead, the carbons from acetyl CoA\text{CoA}CoAC, o, A are initially incorporated into the intermediates of the cycle and are released as carbon dioxide only during later turns. After enough turns, all the carbon atoms from the acetyl group of acetyl CoA\text{CoA}CoAC, o, A will be released as carbon dioxide.
Where’s all the ATP\text{ATP}ATPA, T, P?
You may be thinking that the ATP\text{ATP}ATPA, T, P output of the citric acid cycle seems pretty unimpressive. All that work for just one ATP\text{ATP}ATPA, T, P or GTP\text{GTP}GTPG, T, P?
It’s true that the citric acid cycle doesn’t produce much ATP\text{ATP}ATPA, T, P directly. However, it can make a lot of ATP\text{ATP}ATPA, T, P indirectly, by way of the NADH\text{NADH}NADHN, A, D, H and FADH2\text{FADH}_2FADH2F, A, D, H, start subscript, 2, end subscript it generates. These electron carriers will connect with the last portion of cellular respiration, depositing their electrons into the electron transport chain to drive synthesis of ATP molecules through oxidative phosphorylation.